
David A. Paskewitz and Martin T. Orne
Abstract. Presenting an audible indication of subjects' electroencephalographic alpha activity under conditions of dim ambient illumination led to systematic increases in alpha density, while in total darkness the same procedure did not. These results support the view that feedback training can be clearly demonstrated only when factors leading to a suppression of alpha activity are present in the environment.
Much of the current interest in electroencephalographic (EEG) alpha activity centers around the apparent ability of individuals to learn volitional augmentation of alpha densities in their EEG. Kamiya (1) and Mulholland (2) have independently demonstrated that providing a subject with feedback concerning the presence or absence of his own EEG alpha activity makes it possible for him to alter the amount of such activity seen in the record. In addition, it has been suggested that volitional increases in alpha density lead to changes in subjective mood (3, 4), and it has even been proposed that feedback techniques may be useful as a means of "mapping the subjective space of conscicusness" (5).
The effects of feedback training usually have been demonstrated in one of two ways, both of which contain inherent methodological shortcomings. The first procedure involves comparing periods during which subjects are instructed to maximize alpha density with similar periods when they are instructed to minimize density. Although it is easy to demonstrate differences between periods when subjects are instructed to produce alpha activity and periods when they are instructed to block it, such differences rarely reflect symmetrical increases and decreases. Usually subjects learn to block alpha activity volitionally within one or two trials, and impressive differences with this procedure tend to be the result of unidirectional rather than bidirectional control.
The second procedure involves comparing initial baseline levels with the higher densities seen after training. Such a comparison more appropriately addresses the question whether individuals can learn to augment their level of alpha activity significantly. It then, however, becomes crucial to specify the nature of the baseline level from which increases can be measured. Novelty and apprehension are known to depress the subject's initial alpha density during baseline determination, but as factors such as these dissipate, one tends to see a spontaneously rising baseline. To attempt to follow such a rising baseline, however, through the use of resting periods, interspersed throughout the training sessions as "floating" baselines, does not permit separation of the effects of feedback training from those increases which are due to changes in
arousal or motivation; for the subject, definition of a period as a rest period produces changes in both motivation and arousal.
The present report is intended to help clarify the circumstances under which subjects are able to learn to augment alpha density and shed light on some of the possible mechanisms involved. Specifically, we were concerned with determining whether subjects could learn to significantly exceed the alpha density of their own baseline obtained under optimal circumstances. Because of the known overriding influence of visual stimulation on alpha activity (2), the first study was conducted in total darkness, although to lessen the likelihood of their falling asleep subjects were instructed to keep their eyes open throughout the experiment. Prior to the actual experiment, every effort was made to reassure the subjects and make certain that they were comfortable and understood the equipment and procedures. Nine paid volunteer college students served as subjects, and were recruited for an experiment in "the control of physiological responses." The subjects were given six training sessions, approximately 1 week apart. The monopolar occipital (02) to right mastoid EEG signals were recorded on a Beckman polygraph and filtered with step-function cutoffs at 8 and 12 hz (6). Although the amplitude levels necessary to trigger the feedback stimulus were set in discrete 5-µv steps from 10 to 25 µv for a given subject, depending on initial levels of alpha density, the data used in this report were collected by using an invariant threshold of 15 µv for all subjects. Each session consisted of (i) two 3-minute baseline periods, (ii) a 5-minute orientation period during which feedback was provided and subjects were encouraged to experiment, and (iii) ten trials of 2 minutes each, alternated with 1-minute rest periods. The presence of alpha activity raised the pitch of a feedback stimulus tone from 280 to 360 cycles per second, and activated a clock, the output of which recorded alpha density in seconds per minute. The subjects were instructed that during the trials they should keep the higher (alpha) tone on as much of the time as possible. The absence of any tone served to signal a rest period. At the beginning of each rest period a digital display was illuminated briefly to give the subject a cumulative total of his alpha activity for each trial, and the subject was required to repeat the total out loud.
360
361
An examination of the results for the first session, as shown in Fig. 1, reveals that although subjects' initially high alpha activity was decreased markedly by opening their eyes in total darkness, they had recovered much of this drop by the middle of the initial 5-minute orientation period when the feedback tones were first available to the subjects. These increases occurred within 2 or 3 minutes, without volitional effort. Whether they represent true learning or adaptation is unclear, but the rapidity of these effects is different from what is usually described in alpha feedback training. It furthermore appears that subjects had approached their asymptotic density during the initial orientation period. The subjects exhibit a nonsignificant maximum further increase of 7.2 percent from their highest single minute during the orientation period to the highest density reached during any of the ten trials where subjects were instructed to maximize alpha density (t = 1.81, P > .10). Although in the group data the resting levels tended to be below trial levels, these differences were not significant.
Within the first session, subjects' highest trial densities were significantly lower than their initial eyes-closed baseline densities (t = 2.46, P <.05). When both trial and resting averages are examined for all six sessions, with an analysis of variance repeated measures design, none of the differences are significant (trials: F=0.19, P>.20; rest: F = 0.05, P > .20), and the largest difference between any two trial averages is only about 4 seconds of alpha activity in a minute. The trial average for the sixth session was not greater than the level of alpha density reached during the third minute of the orientation feedback period (t = 0.35, P > .20).
These findings contrast with data reported by others, which imply large increases in alpha density as a consequence of alpha feedback training. We had previously proposed that subjects in the alpha feedback situation are involved in overcoming factors which normally exert an inhibitory influence on alpha activity (7), and we reasoned, therefore, that the lack of consistent increases beyond 2 minutes in our subjects when run in total darkness may well have stemmed from the relative absence of inhibitory stimuli in the situation, especially visual stimuli which Mulholland and Peper have emphasized as primary influences on alpha activity (2, 8). Another group of seven subjects was therefore placed in the same training situation with auditory feedback and exposed to the identical procedure used with the previous group, adding only the presence of dim ambient light.
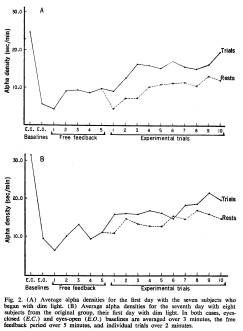
The results for these subjects are shown in Fig. 2A. With this new group, the same drop from eyes-closed to eyes-open baseline is evident. The recovery during the free orientation period preceding the trials is not, however, as great as that seen in darkness. Unlike the earlier group, this second group tested in ambient light exhibited alpha activity during trial periods which was significantly higher than that seen during resting periods (t = 4.33, P < .01) and, furthermore, densities increased beyond the orientation period, as evidenced in a significant 96.8 percent increase in densities from the highest minute during the free orientation period to a subject's highest trial density (t = 4.06, P < .01).
Figure 2A resembles some of the reported findings (1, 3, 9), showing clear-cut increases in alpha density with training. These data would, of course, be even more impressive if the initial eyes-closed baseline were not included in the graph. Most previous reports on the effects of alpha feedback have shown little in the way of such a baseline, and have not considered the possible effects of ambient light.
The significance of the presence of light is seen most clearly by comparing the absence of continuing augmentation in the original group of subjects, run for six sessions in total darkness, with the same group's response during a single session in the presence of dim ambient light. Eight of the original nine subjects were presented with the identical audi-
362
tory feedback procedure they had previously experienced, with only dim ambient light added. The results for this original group, during their seventh session, the first session with light in the room, are shown in Fig. 2B. Recovery from the initial drop takes place slowly, and a significant increase of 55.7 percent was evident from subjects' highest minute during the orientation period to their highest trial density (t = 3.04, P < .02). The difference between trial and resting averages was significant (t = 2.47, P < .05).
Tests between the results of the first session in total darkness and the subsequent session with dim ambient light indicate that the session with light for those same subjects was significantly different, both in trial averages (t = 9.11, P < .001) and resting averages (t = 5.57, P <.001), from their performance in darkness. The session with light for the independent group of subjects also differed significantly from the session in total darkness, both in trial averages (t = 1.96, P < .10) and resting averages (t = 2.64, P < .05). No significant differences exist between the two sessions with light.
Though there may have been some effects of alpha feedback training for six sessions in the dark, the data appear to indicate that the response of the two groups of subjects to alpha feedback training is more alike in the presence of light than the responses of one of those groups with and without the presence of dim light.
The significant differences in the relative level of resting alpha densities between light and dark conditions are probably a function of the fact that light allows the subject to engage in visual scanning during the resting periods, in contrast to the limited scanning possible in total darkness. These results help to explain the discrepancy between Kamiya's findings (1), which showed resting periods at about the same level as during the trial periods, and Brown's results (3), where subjects exhibited resting levels significantly lower than those seen during feedback. Kamiya used an auditory feedback stimulus, and usually had subjects close their eyes. Brown, in contrast, used a visual stimulus, a procedure which necessitated the subjects' keeping their eyes open in a lighted room.
The popular view that alpha feedback training somehow permits individuals to increase alpha densities significantly above those obtained during a comfortable baseline with eyes closed in a nondrowsy, relaxed state cannot be supported by our data. Indeed, despite many training sessions, we have failed to note any individuals whose capacities for producing alpha activity were altered by the feedback procedure. What does seem clear, however, is that alpha feedback training can lead to large and significant changes in alpha densities when conditions have lowered alpha density below the levels seen spontaneously under optimal conditions. Subjects can acquire volitional control over alpha activity only under conditions which normally lead to decreased densities. Thus, during the initial orientation in the dark, when subjects experiment, but are not especially set to increase alpha density, they achieve a level approximating the level reached with additional training. In the light condition, however, there are marked volitional increases. In spite of these increases, however, in the three procedures reported here, as well as during a number of other procedures run in our laboratory, we have not seen alpha densities beyond an individual's initially demonstrated normal physiological range.
While ambient light is sufficient in the present studies to act as a suppressing stimulus, it is likely that other stimuli such as anxiety or physical stress may, in some circumstances, also lead to suppression which persists. Although no increases in alpha density are seen when the situation presents few suppressing stimuli, alpha feedback training may enable a subject to overcome suppressing effects when they are present. The subjective experiences reported to be associated with alpha feedback training (3, 4) may be understood as a consequence of acquiring skill in disregarding stimuli in the external and, perhaps, internal environment which would ordinarily inhibit alpha activity. Increased alpha densities, then, may best he viewed not as an end in themselves, but as one convenient index of a subject's ability to acquire this skill.
Unit for Experimental Psychiatry, Institute of the Pennsylvania Hospital, and University of Pennsylvania, 111 North 49 Street, Philadelphia 19139
References and Notes
1. J. Kamiya, in Altered States of Consciousness: A Book of Readings, C. T. Tart, Ed. (Wiley, New York, 1969), p. 507.
2. T. Mulholland, Activ. Nerv. Super. 10, 410 (1968).
3. B. B. Brown, Psychophysiology 6, 442 (1970).
4. D. P. Nowlis and J. Kamiya, ibid., p. 476.
5. J. Stoyva and J. Kamiya, Psychol. Rev. 75, 192 (1968).
6. D. A. Paskewitz, Psychophysiology 8, 107 (1971).
7. ______, J. J. Lynch, M. T. Orne, J. G. Costello, ibid. 6, 637 (1970); J. J. Lynch and D. A. Paskewitz, J. Nerv. Ment. Dis. 153, 205 (1971).
8. T. Mulholland and E. Peper, Psychophysiology 8, 556 (1971).
9. J. T. Hart, ibid. 4, 506 (1968).
10. We thank our colleagues H. D. Cohen. M. R. Cook, F. J. Evans, C. Graham, R. Hufgard, F. Mural, E. C. Orne, S. F. Pilato, J. W. Powell, L. L. Pyles, and D. S. Roby for comments, criticisms, and technical assistance during the course of this research and the preparation of this report. The research reported here was supported in part by the Advanced Projects Agency of the Department of Defense and was monitored by the Office of Naval Research under contract N00014-70-C-0350 to the San Diego State College Foundation. Reprint requests should be sent to M.T.O.
15 January 1973; revised 18 April 1973